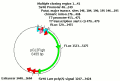
IRESite record type: plasmid_with_promoter_and_putative_IRES_without_translational_characterization
The shape of the nucleic acid molecule translated: linear
The quality of the mRNA/+RNA sequence: possibly_wrong
The mRNA/+RNA description:
Putative in vivo spliced transcript of pIRESneo plasmid from CLONTECH with synthetic intron spliced out. The
intron boundaries do not conform the GT-AG consensus.
The mRNA/+RNA sequence represented in the +DNA notation:
Warning: mRNA sequence when devoid of trailing 'A's is still not a substring of the plasmid sequence. Is it because an intron is spliced out? Stay calm then. :-)
Credibility of mRNA sequence: reverse_engineered_fragment_and_the_rest_is_a_guess
The description of the protein encoded in this ORF: aminoglycoside-3'-O-phosphotransferase
The translational frameshift (ribosome slippage) involved: 0
The ribosome read-through involved: no
The alternative forms of this protein occur by the alternative initiation of translation: not tested
The ORF absolute position (the base range includes START and STOP codons or their equivalents): 778-1581
Remarks:
The intron region annotated in Clontech's PT3043-5.pdf does not follow the GT...AG intron consensus sequence.
The restriction site positions shown in the picture seem to be positions of the next base following
immediately after the place of cleavage. The sequence used here is the one from Clontech's website in the file
PT3043-SEQ.pdf from Mar 31 1998. Unfortunately, the document refers to GenBank sequence U89673 from 1997 which
is shorter by 4 bases after the IVS (intron) sequence while having several base
mismatches/insertions/deletions. We hope the sequence from 1998 is more correct. Let us note the sequence
deposited in GenBank contains immediately after the intron region PvuI site due to those extra 4 bases.
The mRNA sequence considered here is probably partly wrong due to the mis-annotated intron region by Clontech
and starts from the putative CMV immediate early promoter transcription start site and continues up to the
very end of BGH poly(A) signal.
The IRES absolute position (the range includes START and STOP codons or their equivalents): 169-755
How IRES boundaries were determined: experimentally_determined
The sequence of IRES region aligned to its secondary structure (if available):
Remarks:
We consider here the IRES sequence should map against the EMCV Rueckert strain and therefore is annotated
shorter on the left side because 'CG' bases do not align the poly(C) tract of the virus. By aligning the
plasmid against genomic sequence of the Rueckert strain (GenBank accession M81861) the region annotated by
IRESite is extended downstream up to the second ATG codon within EMCV polyprotein CDS region, so the IRES
spans +12nt into the ORF and effectively translation initiates at ATG[13-15] while the ATG[1-3] codon has been
mutated.
Alignment of the IRES cloned into pIRESneo against genomic Ruckert strain sequence is shown below:
pIRESneo ------------------------------------------------------------
M81861 TTGAAAGCCGGGGGTGGGAGATCCGGATTGCCAGTCTGCTCGATATCGCAGGCTGGGTCC 60
pIRESneo ------------------------------------------------------------
M81861 GTGACTACCCACTCCCCCTTTCAACGTGAAGGCTACGATAGTGCCAGGGCGGGTACTGCC 120
pIRESneo ------------------------------------------------------------
M81861 GTAAGTGCCACCCCAAAATAACAACAGACCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCC 180
pIRESneo ------------------------------------------------------------
M81861 CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCC 240
pIRESneo -------------------CCCCTCTCCCTCCCCCCCCCCTAACGTTACTGGCCGAAGCC 41
M81861 CCCCCCCCCCCCCCCCCCCCCCCTCTCCCTCCCCCCCCCCTAACGTTACTGGCCGAAGCC 300
*****************************************
pIRESneo GCTTGGAATAAGGCCGGTGTGCGTTTGTCTATATGTGATTTTCCACCATATTGCCGTCTT 101
M81861 GCTTGGAATAAGGCCGGTGTGCGTTTGTCTATATGTTATTTTCCACCATATTGCCGTCTT 360
************************************ ***********************
pIRESneo TTGGCAATGTGAGGGCCCGGAAACCTGGCCCTGTCTTCTTGACGAGCATTCCTAGGGGTC 161
M81861 TTGGCAATGTGAGGGCCCGGAAACCTGGCCCTGTCTTCTTGACGAGCATTCCTAGGGGTC 420
************************************************************
pIRESneo TTTCCCCTCTCGCCAAAGGAATGCAAGGTCTGTTGAATGTCGTGAAGGAAGCAGTTCCTC 221
M81861 TTTCCCCTCTCGCCAAAGGAATGCAAGGTCTGTTGAATGTCGTGAAGGAAGCAGTTCCTC 480
************************************************************
pIRESneo TGGAAGCTTCTTGAAGACAAACAACGTCTGTAGCGACCCTTTGCAGGCAGCGGAACCCCC 281
M81861 TGGAAGCTTCTTGAAGACAAACAACGTCTGTAGCGACCCTTTGCAGGCAGCGGAACCCCC 540
************************************************************
pIRESneo CACCTGGCGACAGGTGCCTCTGCGGCCAAAAGCCACGTGTATAAGATACACCTGCAAAGG 341
M81861 CACCTGGCGACAGGTGCCTCTGCGGCCAAAAGCCACGTGTATAAGATACACCTGCAAAGG 600
************************************************************
pIRESneo CGGCACAACCCCAGTGCCACGTTGTGAGTTGGATAGTTGTGGAAAGAGTCAAATGGCTCT 401
M81861 CGGCACAACCCCAGTGCCACGTTGTGAGTTGGATAGTTGTGGAAAGAGTCAAATGGCTCT 660
************************************************************
pIRESneo CCTCAAGCGTATTCAACAAGGGGCTGAAGGATGCCCAGAAGGTACCCCATTGTATGGGAT 461
M81861 CCTCAAGCGTATTCAACAAGGGGCTGAAGGATGCCCAGAAGGTACCCCATTGTATGGGAT 720
************************************************************
pIRESneo CTGATCTGGGGCCTCGGTGCACATGCTTTACATGTGTTTAGTCGAGGTTAAAAAAACGTC 521
M81861 CTGATCTGGGGCCTCGGTGCACATGCTTTACATGTGTTTAGTCGAGGTTAAAAAA-CGTC 779
******************************************************* ****
pIRESneo TAGGCCCCCCGAACCACGGGGACGTGGTTTTCCTTTGAAAAACACGATGATAAGCTTGCC 581
M81861 TAGGCCCCCCGAACCACGGGGACGTGGTTTTCCTTTGAAAAACACGATGATAATATGGCC 839
***************************************************** * ***
pIRESneo ACAACC 587
M81861 ACAACC 845
******